UNIDAD 3: EL CORAZÓN
CAPÍTULO 9: MÚSCULO CARDÍACO
El corazón esta formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre a través de la circulación sistémica que aporta flujo sanguíneo a los demás órganos y tejidos del cuerpo. A su vez, cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo.
Cada una de las aurículas es una bomba débil de cebado del ventrículo, que contribuye a transportar sangre hacia el ventrículo correspondiente. Los ventrículos después después aportan la principal fuerza del bombeo que impulsa la sangre: 1)hacia la circulación pulmonar por el ventrículo derecho, o 2) hacia la circulación sistémica por el ventrículo izquierdo.
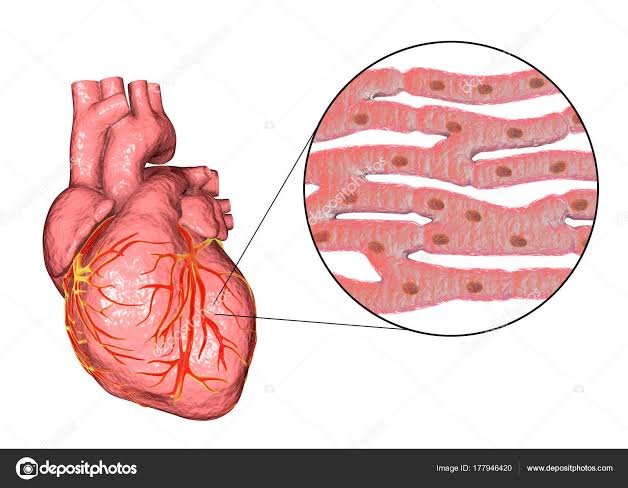
FISIOLOGÍA DEL MÚSCULO CARDÍACO
El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto porque la duración de la contracción es mucho mayor. No obstante, las fibras especializadas de excitación y de conducción del corazón se contraen solo débilmente porque contienen pocas fibrillas contráctiles; en cambio, presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción o conducción de los potenciales de acción por todo el corazón, formando así un sistema excitador que controla el latido rítmico cardíaco.
EL MÚSCULO CARDÍACO ES UN SINCITIO.
Las zonas oscuras que atraviesan las fibras musculares cardíacas se denominan discos intercalados; realmente son membranas celulares que separan las células musculares cardíacas individuales entre sí. Es decir, las fibras musculares cardíacas están formadas por muchas células individuales conec
tadas entre sí en serie y en paralelo. En cada uno de los discos intercalados las membranas celulares se fusionan entre sí para formar uniones «comunicantes» (en hendidura) permeables que permiten una rápida difusión.
Desde un punto de vista funcional los iones se mueven con facilidad en el líquido intracelular a lo largo del eje longitudinal de las fibras musculares cardíacas, de modo que los potenciales de acción viajan fácilmente desde una célula muscular cardíaca a la siguiente, a través de los discos intercalados. Por tanto, el músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una célula se excita el potencial de acción se propaga rápidamente a todas.
El corazón realmente está formado por dos sincitios: el sincitio auricular, que forma las paredes de las dos aurículas, y el sincitio ventricular, que forma las paredes de los dos ventrículos. Las aurículas están separadas de los ventrículos por tejido fibroso que rodea las aberturas de las válvulas aurículoventriculares (AV) entre las aurículas y los ventrículos. Normalmente los potenciales no se conducen desde el sincitio auricular hacia el sincitio ventricular directamente a través de este tejido fibroso. Por el contrario, solo son conducidos por medio de un sistema de conducción especializado denominado haz AV, que es un fascículo de fibras de conducción de varios milímetros de diámetro.
POTENCIALES DE ACCIÓN EN EL MÚSCULO CARDÍACO.
El potencial de acción que se registra en una fibra muscular ventricular, que se muestra en la, es en promedio de aproximadamente 105 mV, lo que significa que el potencial intracelular aumenta desde un valor muy negativo, de aproximadamente –85 mV, entre los latidos hasta un valor ligeramente positivo, de aproximadamente +20 mV, durante cada latido. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una meseta, seguida al final de la meseta de una repolarización súbita. La presencia de esta meseta del potencial de acción ha- ce que la contracción ventricular dure hasta 15 veces más en el músculo cardíaco que en el músculo esquelético.
¿QUÉ PRODUCE EL POTENCIAL DE ACCIÓN PROLONGADO Y LA MESETA?
En el músculo cardíaco, el potencial de acción está producido por la apertura de dos tipos de canales: 1) los mismos canales rápidos de sodio activados por el voltaje que en el músculo esquelético y 2) otra población totalmente distinta de canales de calcio de tipo L (canales lentos de calcio), que también se denominan canales de calcio-sodio.
RESUMEN DE LAS FASES DEL POTENCIAL DE ACCIÓN DEL MÚSCULO CARDÍACO.
Fase 0 (despolarización), los canales de sodio rápidos se abren. Cuando la célula cardíaca es estimulada y se despolariza, el potencial de membrana se hace más positivo. Los canales de sodio activados por el voltaje (canales de sodio rápidos) se abren y permiten que el sodio circule rápidamente hacia el interior de la célula y la despolarice. El potencial de membrana alcanza +20 mV aproximadamente antes de que los canales de sodio se cierren.
Fase 1 (repolarización inicial), los canales de sodio rápidos se cierran. Los canales de sodio se cierran, la célula empieza a repolarizarse y los iones potasio salen de la célula a través de los canales de potasio.
Fase 2 (meseta), los canales de calcio se abren y los canales de potasio rápidos se cierran. Tiene lugar una breve repolarización inicial y el potencial de acción alcanza una meseta como consecuencia de: 1) una mayor permeabilidad a los iones calcio, y 2) la disminución de la permeabilidad a los iones potasio.
Fase 3 (repolarización rápida), los canales de calcio se cierran y los canales de potasio lentos se abren. El cierre de los canales iónicos de calcio y el aumento de la permeabilidad a los iones potasio, que permiten que los iones potasio salgan rápidamente de la célula, pone fin a la meseta y devuelve el potencial de membrana de la célula a su nivel de reposo.
Fase 4 (potencial de membrana de reposo) con valor medio aproximado de -90 mV.
VELOCIDAD DE LA CONDUCCIÓN DE LAS SEÑALES EN EL MÚSCULO CARDÍACO.
La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de unos 0,3 a 0,5 m/s, o aproximadamente 1/250 de la velocidad en las fibras nerviosas grandes y en torno a 1/10 de la velocidad en las fibras musculares esqueléticas. La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras de Purkinje, es de hasta 4 m/s.
PERIODO REFRACTARIO DEL MÚSCULO CARDÍACO
El período refractario normal del ventrículo es de 0,25 a 0,30 s, que es aproximadamente la duración del potencial de acción en meseta prolongado, Hay un período relativo refractario adicional de aproximadamente 0,05 s, durante el cual es más difícil de lo normal excitar el músculo pero, sin embargo, se puede excitar con una señal excitadora muy intensa.
ACOPLAMIENTO EXCITACIÓN-CONTRACCIÓN: FUNCIÓN DE LOS IONES CALCIO Y DE LOS TÚBULOS TRANSVERSOS.
El término «acoplamiento excitación-contracción» se refiere al mecanismo mediante el cual el potencial de acción hace que las miofibrillas del músculo se contraigan. Una vez más hay diferencias en este mecanismo en el músculo cardíaco que tienen efectos importantes sobre las características de su contracción. Al igual que en el músculo esquelético, cuando un potencial de acción pasa sobre la membrana del músculo cardíaco el potencial de acción se propaga hacia el interior de la fibra muscular cardíaca a lo largo de las membranas de los túbulos transversos (T). Los potenciales de acción de los túbulos T, a su vez, actúan sobre las membranas de los túbulos sarcoplásmicos longitudinales para producir la liberación de iones calcio hacia el sarcoplasma muscular desde el retículo sarcoplásmico. En algunas milésimas de segundo más estos iones calcio difunden hacia las miofibrillas y catalizan las reacciones químicas que favorecen el deslizamiento de los filamentos de actina y de miosina entre sí, lo que da lugar a la contracción muscular.
El calcio que entra en la célula activa después los canales de liberación de calcio, también denominados canales de receptor de rianodina, en la membrana del retículo sarcoplásmico, para activar la liberación de calcio en el sarcoplasma.
CICLO CARDÍACO.
Cada ciclo es iniciado por la generación espontánea de un potencial de acción en el nódulo sinusal. Este nódulo está localizado en la pared superolateral de la aurícula derecha, cerca del orificio de la vena cava superior, y el potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos. Las aurículas actúan como bombas de cebado para los ventrículos, y los ventrículos a su vez proporcionan la principal fuente de potencia para mover la sangre a través del sistema vascular del cuerpo.
DIÁSTOLE Y SÍSTOLE.
El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole. La duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca. Por ejemplo, si la frecuencia cardíaca es de 72 latidos/min, la duración del ciclo cardíaco es de 1/72 min/latido, aproximadamente 0,0139 min por latido, o 0,833 s por latido.
RELACIÓN DEL ELECTROCARDIOGRAMA CON EL CICLO CARDÍACO.
Son los voltajes eléctricos que genera el corazón, y son registrados mediante el electrocardiógrafo desde la superficie del cuerpo. La onda P está producida por la propagación de la despolarización en las aurículas, y es seguida por la contracción auricular, que produce una ligera elevación de la curva de presión auricular inmediatamente después de la onda P electrocardiográfica. Aproximadamente 0,16 s después del inicio de la onda P, las ondas QRS aparecen como consecuencia de la despolarización eléctrica de los ventrículos, que inicia la contracción de los ventrículos y hace que comience a elevarse la presión ventricular. Por tanto, el complejo QRS comienza un poco antes del inicio de la sístole ventricular. Finalmente, la onda T ventricular representa la fase de repolarización de los ventrículos, cuando las fibras del músculo ventricular comienzan a relajarse. Por tanto, la onda T se produce un poco antes del final de la contracción ventricular.
FUNCIÓN DE LAS AURÍCULAS COMO BOMBAS DE CEBADO PARA LOS VENTRÍCULOS
La sangre normalmente fluye de forma continua desde las grandes ventas hacia las aurículas; aproximadamente el 80% de la sangre fluye directamente a través de las aurículas hacia los ventrículos incluso antes de que se contraigan las aurículas. Después, la contracción auricular habitualmente produce un llenado de un 20% adicional de los ventrículos. Por tanto, las aurículas actúan como bombas de cebado que aumentan la eficacia del bombeo ventricular hasta un 20%. Sin embargo, el corazón puede seguir funcionando en la mayor parte de las condiciones incluso sin esta eficacia de un 20% adicional porque normalmente tiene la capacidad de bombear entre el 300 y el 400% más de sangre de la que necesita el cuerpo en reposo. Por tanto, cuando las aurículas dejan de funcionar es poco probable que se observe esta diferencia salvo que la persona haga un esfuerzo; en este caso de manera ocasional aparecen síntomas agudos de insuficiencia cardíaca, especialmente disnea.
DESBORDAMIENTO DE LOS VENTRÍCULOS DURANTE LA SÍSTOLE
Es el llamado período de contracción isovolumétrica o isométrica, lo que quiere decir que se produce aumento de la tensión en el músculo cardíaco, pero con un acortamiento escaso o nulo de las fibras musculares.
PERIODO DE EYECCIÓN
Cuando la presión ventricular izquierda aumenta ligeramente por encima de 80 mmHg (y la presión ventricular derecha ligeramente por encima de 8 mmHg), las presiones ventriculares abren las válvulas semilunares. Inmediatamente comienza a salir la sangre de los ventrículos. Aproximadamente el 60% de la sangre del ventrículo al final de la diástole es expulsada durante la sístole; en torno al 70% de esta porción es expulsado durante el primer tercio del período de eyección y el 30% restante del vaciado se produce durante los dos tercios siguientes. El primer tercio se denomina período de eyección rápida y los dos tercios finales período de eyección lenta.
PERIODO DE RELAJACIÓN ISOVOLUMÉTRICA (ISOMÉTRICA)
Las presiones elevadas de las grandes arterias distendidas que se acaban de llenar con la sangre que procede de los ventrículos que se han contraído empujan inmediatamente la sangre de nuevo hacia los ventrículos, lo que cierra súbitamente las válvulas aórtica y pulmonar. Durante otros 0,03 a 0,06 s el músculo cardíaco sigue relajándose, aun cuando no se modifica el volumen ventricular, dando lugar al período de relajación isovolumétrica o isométrica.
LAS VÁLVULAS CARDÍACAS EVITAN EL FLUJO INVERSO DE LA SANGRE DURANTE LA SÍSTOLE.
VÁLVULAS AURÍCULOVENTRICULARES.
Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (es decir, las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole. Por motivos anatómicos, las válvulas AV, que están formadas por una película delgada, casi no precisan ningún flujo retrógrado para cerrarse, mientras que las válvulas semilunares, que son mucho más fuertes, precisan un flujo retrógrado bastante rápido durante algunos milisegundos.
VÁLVULAS AÓRTICA Y DE LA ARTERIA PULMONAR.
Las válvulas semilunares aórtica y pulmonar funcionan de una manera bastante distinta de las válvulas AV. Primero, las elevadas presiones de las arterias al final de la sístole hacen que las válvulas semilunares se cierren súbitamente, a diferencia del cierre mucho más suave de las válvulas AV. Segundo, debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores.
CURVA DE PRESIÓN AÓRTICA.
La entrada de sangre en las arterias durante la sístole hace que sus paredes se distiendan y que la presión aumente hasta aproximadamente 120 mmHg. Se produce una incisura en la curva de presión aórtica cuando se cierra la válvula aórtica; está producida por un corto período de flujo retrógrado de sangre inmediatamente antes del cierre de la válvula, seguido por la interrupción súbita del flujo retrógrado.
Antes de que se contraiga de nuevo el ventrículo, la presión aórtica habitualmente ha disminuido hasta aproximadamente 80 mmHg (presión diastólica), que es dos tercios de la presión máxima de 120 mmHg (presión sistólica) que se produce en la aorta durante la contracción ventricular. Las curvas de presión del ventrículo derecho y de la arteria pulmonar son similares a las de la aorta, excepto que las presiones tienen una magnitud de solo aproximadamente 1/6.
<> DURANTE EL CICLO CARDÍACO: TRABAJO CARDÍACO.
Fase I: período de llenado. La fase I del diagrama volumen-presión comienza a un volumen ventricular de aproximadamente 50 ml y una presión diastólica próxima a 2 a 3 mmHg. La cantidad de sangre que queda en el ventrículo después del latido previo, 50 ml, se denomina volumen telesistólico.
Fase II: período de contracción isovolumétrica. Durante la contracción isovolumétrica el volumen del ventrículo no se modifica porque todas las válvulas están cerradas. Sin embargo, la presión en el interior del ventrículo aumenta hasta igualarse a la presión que hay en la aorta, hasta un valor de presión de aproximadamente 80 mmHg.
Fase III: período de eyección. Durante la eyección la presión sistólica aumenta incluso más debido a una contracción aún más intensa del ventrículo. Al mismo tiempo, el volumen del ventrículo disminuye porque la válvula aórtica ya se ha abierto y la sangre sale del ventrículo hacia la aorta.
Fase IV: período de relajación isovolumétrica. Al final del período de eyección se cierra la válvula aórtica, y la presión ventricular disminuye de nuevo hasta el nivel de la presión diastólica. La línea marcada como «IV» refleja esta disminución de la presión intraventricular sin cambios de volumen. Así, el ventrículo recupera su valor inicial, en el que quedan aproximadamente 50 ml de sangre en el ventrículo y la presión auricular es de aproximadamente de 2 a 3 mmHg. El área que encierra este diagrama volumen-presión funcional (la zona sombreada denominada TE) representa el trabajo cardíaco externo neto del ventrículo durante su ciclo de contracción.
CONCEPTOS DE PRECARGA Y POSCARGA.
Cuando se evalúan las propiedades contráctiles del músculo es importante especificar el grado de tensión del músculo cuando comienza a contraerse, que se denomina precarga y especificar la carga contra la que el músculo ejerce su fuerza contráctil, que se denomina poscarga. La precarga es la presión telediastólica cuando el ventrículo ya se ha llenado. La poscarga del ventrículo es la presión de la aorta que sale del ventrículo. La importancia de los conceptos de precarga y poscarga es que en muchos estados funcionales anormales del corazón o de la circulación, la presión durante el llenado del ventrículo (la precarga), la presión arterial contra la que se debe contraer el ventrículo (la poscarga) o ambas están alteradas con respecto a su situación normal en un grado importante.
EFECTO DE LA TEMPERATURA SOBRE LA FUNCIÓN CARDÍACA.
El aumento de la temperatura corporal, como ocurre durante la fiebre, produce un gran aumento de la frecuencia cardiaca, a veces hasta el doble del valor normal. El descenso de la temperatura produce una gran disminución de la frecuencia cardiaca, que puede disminuir hasta solo algunos latidos por minuto cuando una persona esta cerca de la muerte por hipotermia en el intervalo de temperatura corporal de 16 a 21 grados centígrados.
Brandon nazario vazquez balanzar
