Contracción del músculo esquelético
En este tema veremos los diferentes músculos y los tipos de contracciones que realizan y como son sus disposiciones. Aproximadamente el 40% del cuerpo es músculo esquelético, y tal vez otro 10% es músculo liso y cardíaco. Algunos de los principios básicos de la contracción se aplican también a los diferentes tipos de músculos.
Anatomía fisiológica del músculo esquelético
Fibras del músculo esquelético
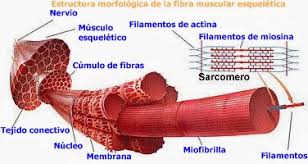
Todos los músculos esqueléticos están formados por numerosas fibras cuyo diámetro varía entre 10 y 80 μm. Cada una de estas fibras está formada por subunidades cada vez más pequeñas.
En la mayoría de los músculos esqueléticos, las fibras se extienden a lo largo de toda la longitud del músculo. Todas las fibras, excepto alrededor de un 2%, habitualmente están inervadas por una sola terminación nerviosa localizada cerca del punto medio de la fibra.
El sarcolema es una fina membrana que envuelve a una fibra musculoesquelética
El
sarcolema está formado por una membrana celular verdadera, denominada membrana plasmática, y una cubierta externa formada por una capa delgada de material
polisacárido que contiene numerosas fibrillas delgadas de colágeno. En cada uno
de los dos extremos de la fibra muscular la
capa superficial del sarcolema se fusiona con una fibra tendinosa. Las fibras
tendinosas a su vez se agrupan en haces para formar los tendones musculares,
que después insertan los músculos en los huesos.
Las miofibrillas están formadas por filamentos de actina y miosina
Cada fibra muscular contiene varios cientos a varios miles de miofibrillas Cada miofibrilla está formada por aproximadamente 1.500 filamentos de miosina y 3.000 filamentos de actina adyacentes entre sí, que son grandes moléculas proteicas polimerizadas responsables de la contracción muscular real.
los
filamentos de miosina y de actina se interdigitan parcialmente
y de esta manera hacen que las miofibrillas tengan bandas claras y oscuras
alternas.
Las bandas claras contienen solo filamentos de actina y se denominan bandas I porque son isótropas a la luz polarizada. Las bandas oscuras contienen filamentos de miosina, así como los extremos de los filamentos de actina en el punto en el que se superponen con la miosina, y se denominan bandas A porque son anisótropas a la luz polarizada.
Las moléculas filamentosas de titina mantienen en su lugar los filamentos de miosina y actina
La relación
de yuxtaposición entre los filamentos de miosina y de actina se mantiene por
medio de un gran número de moléculas filamentosas de una proteína denominada titina. Cada molécula de titina tiene un peso molecular de aproximadamente
3 millones, lo que hace que sea una de las
mayores moléculas proteicas del cuerpo. Además, como es filamentosa, es muy elástica. Estas moléculas elásticas
de titina actúan como armazón que mantiene en su posición los filamentos de
miosina y de actina, de modo que funcione la maquinaria contráctil del
sarcómero. Un extremo de la
molécula de titina es elástico y está unido al disco Z; para actuar a modo de
muelle y con una longitud que cambia según el sarcómero se contrae y se relaja.
La otra parte de la molécula de titina la une al grueso filamento de miosina.
La molécula de titina también parece actuar como molde para la formación
inicial de porciones de los filamentos contráctiles del sarcómero,
especialmente los filamentos de miosina.
El
sarcoplasma es el fluido intracelular entre las miofibrillas
Las muchas
miofibrillas de cada fibra muscular están yuxtapuestas suspendidas en la fibra
muscular. Los espacios entre las miofibrillas están llenos de un líquido
intracelular denominado sarcoplasma,
que contiene grandes cantidades de potasio, magnesio y fosfato,
además de múltiples enzimas proteicas. También hay muchas mitocondrias que están dispuestas
paralelas a las miofibrillas. Estas mitocondrias proporcionan a las
miofibrillas en contracción grandes cantidades de energía en forma
de trifosfato de adenosina (ATP), que es formado por las mitocondrias.
El retículo
sarcoplásmico es un retículo endoplásmico especializado de músculoesquelético
En el sarcoplasma que rodea a las miofibrillas de todas las fibras musculares también hay un extenso retículo denominado retículo sarcoplásmico. Este retículo tiene una organización especial que es muy importante para regular el almacenamiento, la liberación y la recaptación de calcio y, por tanto, para controlar la contracción muscula. Los tipos de fibras musculares de contracción rápida tienen retículos sarcoplásmicos especialmente extensos.
Mecanismo
general de la contracción muscular
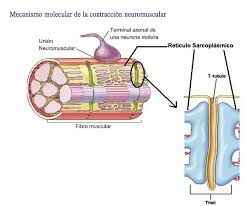
El inicio y la ejecución de la contracción muscular se producen en las siguientes etapas secuenciales:
1. Un potencial de acción viaja a lo largo de una fibra motora hasta sus terminales sobre las fibras musculares.
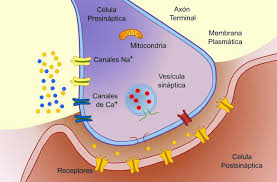
2. En cada terminal, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina.
3. La
acetilcolina actúa en una zona local de la membrana de la fibra muscular para
abrir múltiples canales de cationes «activados por acetilcolina» a través de
moléculas proteicas que flotan en la membrana.
4. La apertura de los canales activados por acetilcolina permite que grandes cantidades de iones sodio difundan hacia el interior de la membrana de la fibra muscular. Esta acción provoca una despolarización local que, a su vez, conduce a la apertura de los canales de sodio activados por el voltaje, que inicia un potencial de acción en la membrana.
5. El potencial de acción viaja a lo largo de la membrana de la fibra muscular de la misma manera que los potenciales de acción viajan a lo largo de las membranas de las fibras nerviosas.
6. El
potencial de acción despolariza la membrana muscular, y buena parte de la
electricidad del potencial de acción fluye a través del centro de la fibra
muscular, donde hace que el retículo sarcoplásmico libere grandes cantidades de
iones calcio que se han almacenado en el interior de este retículo.
7. Los iones calcio inician fuerzas de atracción entre los filamentos de actina y miosina, haciendo que se deslicen unos sobre otros en sentido longitudinal, lo que constituye el proceso contráctil.
8. Después de una fracción de segundo los iones calcio son bombeados de nuevo hacia el retículo sarcoplásmico por una bomba de Ca++ de la membrana y permanecen almacenados en el retículo hasta que llega un nuevo potencial de acción muscular; esta retirada de los iones calcio desde las miofibrillas hace que cese la contracción muscular.
Mecanismo molecular de la contracción muscular
La contracción muscular se produce por un mecanismo de deslizamiento de los filamentos. El estado relajado de un sarcómero (arriba) y su estado contraído (abajo). En el estado relajado, los extremos de los filamentos de actina que se extienden entre dos discos Z sucesivos apenas comienzan a superponerse entre sí. Por el contrario, en el estado contraído estos filamentos de actina han sido traccionados hacia dentro entre los filamentos de miosina, de modo que sus extremos se superponen entre sí en su máxima extensión. Además, los discos Z han sido traccionados por los filamentos de actina hasta los extremos de los filamentos de miosina. Así, la contracción muscular se produce por un mecanismo de deslizamiento de los filamentos.
Los filamentos de miosina están compuestos por multiples moléculas de miosina. Las colas delas moléculas de miosina se agrupan entre sí para formar el cuerpo del filamento, mientras que las cabezas y parte de cada molecula miosina cuelgan por fuera de los lados del cuerpo, formando un brazo que separa la cabeza del cuerpo. Los brazos y las cabezas que protruyen se denominan puentes cruzados. Una característica importante de la cabeza de la miosina es que actua como una enzima adenosina trifosfatasa, lo que le permite escindir el trifosfato de adenosina (ATP) para aportar energía al proceso de la contracción.
Los filamentos de actina están formados por actina, tropomiosina y troponina. Cada uno de los filamentos de actina tiene una longitud de aproximadamente 1um. Las bases de los filamentos de actina se anclan fuertemente en los discos Z mientras que los extremos protruyen en ambas direcciones dentro de los sarcomeros adyacentes para situarse en los espacios que hay entre las moléculas de miosina.
Interacción de un filamento de miosina, dos filamentos de actina y los iones calcio para producir la contracción.
El filamento de actina se inhibe por el complejo troponina-tropomiosina . la activación se estimula por los iones calcio.
Inhibición del complejo troponina-tropomiosina. Los puntos activos del filamento de actina normal del musculo relajado son inhibidos o cubiertos físicamente por el complejo troponina-tropomiosina. En consecuencia, estos puntos no se pueden unir a las cabezas de los filamentos de miosina para producir la contracción hasta que se inhiba asu vez el efecto inhibidor del complejo troponina-tropomiosina.
Activación por los iones de calcio. El efecto inhibidor del complejo troponina-tropomiosina sobre los filamentos de actina se inhibe por la presencia de iones de calcio. Los iones calcio se combinan con la troponina C, provocando que el complejo de troponina tire de a molécula de tropomiosina. Esta acción descubre los puntos activos de la actina, permitiendo que las cabezas de miosina se unan y se produzca la contracción.
La teoría de la cremallera puede explicar cómo interactúan el filamento de actina activado y los puntos cruzados de la miosina para provocar la contracción. Cuando una cabeza se une a un punto activo, la cabeza se desplaza automáticamente hacia el brazo y arrastra con ella al filamento de actina. Este desplazamiento de la cabeza se denomina golpe activo. Inmediatamente después del desplazamiento, la cabeza se separa automáticamente del punto activo. A continuación, la cabeza recupera su dirección extendida. En esta posición, se combina con un nuevo punto activo que está más abajo, a lo largo del filamento de actina. Asi, las cabezas de los puentes cruzados se curvan hacia atrás y hacia adelante y, paso a paso, recorren el filamento de actina, desplazando los extremos de los filamentos de actina hacia el centro del filamento de miosina.
El efecto de la cantidad de superposición de los filamentos de actina y miosina determina la tensión desarrollada por el musculo en contracción.
La potencia de la contracción es máxima cuando la superposición entre los filamentos de actina y los puentes cruzados de los filamentos de miosina es óptima. El musculo no puede desarrollar su fuerza cuando la longitud del sarcomero no fisiológica es muy alta, ya que no existe superposición entre los filamentos de actina y miosina. La tensión aumenta progresivamente a medida que el sarcomero se acorta y los filamentos de actina y miosina comienzan a superponerse. La tensión máxima se mantiene cuando la longitud del sarcomero es de 2 um porque el filamento de actina ha interaccionado con todos los puentes cruzados del filamento de miosina. Con un nuevo acortamiento los extremos de los dos filamentos de actina comienzan a superponerse, reduciendo la tensión muscular. Cuando la longitud del sarcomero disminuye hasta 1.65 um, los dos discos Z del sarcomero se encuentran apoyados en los extremos de los filamentos de miosina y la fuerza de la contracción disminuye drásticamente.
Energética de la contracción muscular
La contracción muscular requiere ATP para realizar tres funciones principales
. La mayor parte del ATP se emplea para activar el mecanismo de cremallera de la contracción muscular.
. El transporte activo de iones calcio hacia el interior del retículo sarcoplasmatico hace que finalice la contracción.
. los iones sodio y potasio se bombean a través de la membrana de la fibra muscular para mantener un entorno iónico adecuado para la propagación de los potenciales de acción.
La energía para la contracción muscular procede de tres fuentes principales. La contracción de ATP en la fibra muscular es suficiente para mantener la contracción completa solo durante 1 a 2 s. El ATP se escinde para formar difosfato de adenosina (ADP) y el ADP se vuelve a fosforilar para formar nuevo ATP. Hay varias fuentes de energía para esta nueva fosforilacion.
La fosfocreatina contiene un enlace fosfato de alta energía similar a los enlaces del ATP pero contiene más energía libre. La energía que se libera de este enlace produce a su vez el enlace de un nuevo ion fosfato inorgánico al ADP para reconstruir el ATP. La energía combinada del ATP y de la fosfocreatina almacenados en el musculo es capaz de producir una concentración muscular máxima solo durante 5 a 8 s.
. La escisión enzimática rápida del glucógeno en acido pirúvico y ácido láctico libera energía que se utiliza para convertir el ADP en ATP. Las reacciones glucoliticas se pueden producir en ausencias de oxígeno. La velocidad de formación de ATP por el proceso glucolitico es aproximadamente 2.5 veces más rápido que la formación de ATP en respuesta a la reacción de los nutrientes celulares con el oxígeno. La glucolisis sola puede mantener una contracción muscular máxima solo durante aproximadamente 1 min.
Características de la contracción de todo el musculo
Las contracciones isométricas no acortan el musculo, mientras que las contracciones isotónicas lo acortan a una tensión constante. La contracción isométrica se produce cuando el musculo no se acorta durante la contracción. Las contracciones isométricas verdaderas no pueden generarse en el cuerpo intacto, ya que los denominados componentes elásticos en serie se estiran durante la contracción, permitiendo un pequeño acortamiento del musculo. Dichos elementos elásticos son los tendones, las terminaciones del sarcolema de las fibras musculares y, quizás, los brazos de los puentes cruzados de miosina.
La contracción isotónica se produce cuando el musculo se acorta, pero la tensión permanece constante. Las características de la contracción isotónica dependen de la carga contra la que el musculo se contrae, además de la inercia de la carga.
Las fibras rápidas están adaptadas para las contracciones musculares potentes, mientras que las lentas están adaptadas para la actividad muscular prolongada. Cada musculo está compuesto por una mezcla de las denominadas fibras musculares rápidas y lentas, con otras fibras intermedias entre estos dos extremos. No obstante, un musculo dado puede tener fibras musculares predominantemente rapidas, mientras que otros pueden tener fibras predominantemente lentas.
Las fibras lentas tipo uno musculo rojo son: más pequeñas tienen capilares sanguíneos más extensos y un número muy elevados de mitocondrias para mantener niveles elevados de metabolismo oxidativos y contienen grandes cantidades de mioglobina, lo que da al musculo lento un aspecto rojizo y el nombre de músculos rojo. El déficit de mioglobina roja en el musculo rápido le da el nombre de musculo blanco.
Las fibras rápidas tipo 2 musculo blanco son: fibras grandes para obtener una gran fuerza de contracción; tiene un retículo sarcoplasmatico extenso para una liberación rápida de iones calcio tiene grandes cantidades de enzimas glucoliticas para la liberación rápida de energía por el proceso glucolitico y tienen una vascularización menos extensa y menos mitocondrias por que el metabolismo oxidativo es secundario.
